Papenfort Kai - Regulating with RNA in Bacteria and Archaea
Here you can read online Papenfort Kai - Regulating with RNA in Bacteria and Archaea full text of the book (entire story) in english for free. Download pdf and epub, get meaning, cover and reviews about this ebook. City: Washington, DC, year: 2019, publisher: ASM Press, genre: Art. Description of the work, (preface) as well as reviews are available. Best literature library LitArk.com created for fans of good reading and offers a wide selection of genres:
Romance novel
Science fiction
Adventure
Detective
Science
History
Home and family
Prose
Art
Politics
Computer
Non-fiction
Religion
Business
Children
Humor
Choose a favorite category and find really read worthwhile books. Enjoy immersion in the world of imagination, feel the emotions of the characters or learn something new for yourself, make an fascinating discovery.

- Book:Regulating with RNA in Bacteria and Archaea
- Author:
- Publisher:ASM Press
- Genre:
- Year:2019
- City:Washington, DC
- Rating:3 / 5
- Favourites:Add to favourites
- Your mark:
Regulating with RNA in Bacteria and Archaea: summary, description and annotation
We offer to read an annotation, description, summary or preface (depends on what the author of the book "Regulating with RNA in Bacteria and Archaea" wrote himself). If you haven't found the necessary information about the book — write in the comments, we will try to find it.
Papenfort Kai: author's other books
Who wrote Regulating with RNA in Bacteria and Archaea? Find out the surname, the name of the author of the book and a list of all author's works by series.










Regulating with RNA in Bacteria and Archaea — read online for free the complete book (whole text) full work
Below is the text of the book, divided by pages. System saving the place of the last page read, allows you to conveniently read the book "Regulating with RNA in Bacteria and Archaea" online for free, without having to search again every time where you left off. Put a bookmark, and you can go to the page where you finished reading at any time.
Font size:
Interval:
Bookmark:
- Section I: RNases and Helicases
- 1. RNase E and the High-Fidelity Orchestration of RNA Metabolism
Katarzyna J. Bandyra and Ben F. Luisi - 2. Enzymes Involved in Posttranscriptional RNA Metabolism in Gram-Negative Bacteria
Bijoy K. Mohanty and Sidney R. Kushner - 3. RNases and Helicases in Gram-Positive Bacteria
Sylvain Durand and Ciarn Condon
- 1. RNase E and the High-Fidelity Orchestration of RNA Metabolism
- Section II: Cis-Acting RNAs
- 4. RNA Thermometers in Bacterial Pathogens
Edmund Loh, Francesco Righetti, Hannes Eichner, Christian Twittenhoff, and Franz Narberhaus - 5. Small Molecule-Binding Riboswitches
Thea S. Lotz and Beatrix Suess - 6. The T-Box Riboswitch: tRNA as an Effector to Modulate Gene Regulation
Kiel D. Kreuzer and Tina M. Henkin - 7. rRNA Mimicry in RNA Regulation of Gene Expression
Michelle M. Meyer - 8. Processive Antitermination
Jonathan R. Goodson and Wade C. Winkler - 9. Genes within Genes in Bacterial Genomes
Sezen Meydan, Nora Vzquez-Laslop, and Alexander S. Mankin - 10. Leaderless mRNAs in the Spotlight: Ancient but Not Outdated!
Heather J. Beck and Isabella Moll
- 4. RNA Thermometers in Bacterial Pathogens
- Section III: Cis-Encoded Base Pairing RNAs
- 11. Type I Toxin-Antitoxin Systems: Regulating Toxin Expression via Shine-Dalgarno Sequence Sequestration and Small RNA Binding
Sara Masachis and Fabien Darfeuille - 12. Widespread Antisense Transcription in Prokaryotes
Jens Georg and Wolfgang R. Hess
- 11. Type I Toxin-Antitoxin Systems: Regulating Toxin Expression via Shine-Dalgarno Sequence Sequestration and Small RNA Binding
- Section IV: Trans-Encoded Base Pairing RNAs
- 13. Small Regulatory RNAs in the Enterobacterial Response to Envelope Damage and Oxidative Stress
Kathrin S. Frhlich and Susan Gottesman - 14. Carbohydrate Utilization in Bacteria: Making the Most Out of Sugars with the Help of Small Regulatory RNAs
Svetlana Durica-Mitic, Yvonne Gpel, and Boris Grke - 15. Small RNAs Involved in Regulation of Nitrogen Metabolism
Daniela Prasse and Ruth A. Schmitz - 16. Bacterial Iron Homeostasis Regulation by sRNAs
Sylvia Chareyre and Pierre Mandin - 17. Small-RNA-Based Regulation of Bacterial Quorum Sensing and Biofilm Formation
Sine Lo Svenningsen - 18. Regulatory RNAs in Virulence and Host-Microbe Interactions
Alexander J. Westermann
- 13. Small Regulatory RNAs in the Enterobacterial Response to Envelope Damage and Oxidative Stress
- Section V: Protein Titration and Scaffolding
- 19. Global Regulation by CsrA and Its RNA Antagonists
Tony Romeo and Paul Babitzke - 20. 6S RNA, a Global Regulator of Transcription
Karen M. Wassarman - 21. Bacterial Y RNAs: Gates, Tethers, and tRNA Mimics
Soyeong Sim and Sandra L. Wolin
- 19. Global Regulation by CsrA and Its RNA Antagonists
- Section VI: General Considerations
- 22. Proteins That Chaperone RNA Regulation
Sarah A. Woodson, Subrata Panja, and Andrew Santiago-Frangos - 23. Epitranscriptomics: RNA Modifications in Bacteria and Archaea
Katharina Hfer and Andres Jschke - 24. RNA Localization in Bacteria
Jingyi Fei and Cynthia M. Sharma - 25. Sponges and Predators in the Small RNA World
Nara Figueroa-Bossi and Lionello Bossi - 26. Bacterial Small RNAs in Mixed Regulatory Networks
Anas Brosse and Maude Guillier - 27. Dual-Function RNAs
Medha Raina, Alisa King, Colleen Bianco, and Carin K. Vanderpool - 28. Origin, Evolution, and Loss of Bacterial Small RNAs
H. Auguste Dutcher and Rahul Raghavan
- 22. Proteins That Chaperone RNA Regulation
- Section VII: Emerging Topics
- 29. Cross-Regulation between Bacteria and Phages at a Posttranscriptional Level
Shoshy Altuvia, Gisela Storz, and Kai Papenfort - 30. Large Noncoding RNAs in Bacteria
Kimberly A. Harris and Ronald R. Breaker - 31. Synthetic Biology of Small RNAs and Riboswitches
Jordan K. Villa, Yichi Su, Lydia M. Contreras, and Ming C. Hammond
- 29. Cross-Regulation between Bacteria and Phages at a Posttranscriptional Level
- Section VIII: Resources
- 32. Functional Transcriptomics for Bacterial Gene Detectives
Blanca M. Perez-Sepulveda and Jay C.D. Hinton - 33. Structure and Interaction Prediction in Prokaryotic RNA Biology
Patrick R. Wright, Martin Mann, and Rolf Backofen
- 32. Functional Transcriptomics for Bacterial Gene Detectives
Katarzyna J. Bandyra
Ben F. Luisi
It may seem surprising that in almost all known life-forms, information-encoding transcripts are actively annihilated. Although at first glance this seems to be a potential waste of resources and loss of information, the anticipated advantages of restricting transcript lifetimes include fast response rates and a capacity to rapidly redirect gene expression pathways. In this way, destroying individual transcripts in a modulated manner might effectively enhance the collective information capacity of the living system. Escherichia coli has proven to be a useful model system to study such processes, and nearly 45 years ago, a hypothetical endoribonuclease was proposed by Apirion as the key missing factor that might account for the observed degradation patterns of mRNA in that bacterium. At the time this hypothesis was formulated, transcript decay in E. coli was best described as a series of endonucleolytic cleavages and subsequent fragment scavenging by 3 exonucleases ().
In the ensuing decades following the discovery of RNase E, more evidence and deeper insights have been gained into the function and importance of the enzyme in RNA metabolism. The data corroborate the numerous roles played by the RNase, including the initiation of turnover for many mRNA species ().
It is important to note that RNase E is not the sole RNase that can initiate turnover in E. coli , as others can catalyze the initial cleavage of mRNAs, including RNase G, RNase P, the double-strand-specific RNase III, and RNases from the toxin/antitoxin families (), implicating a unique and dominating role.
The access of RNase E and other RNases to substrates can be modulated by RNA-binding proteins (). These local structures can be induced or remodeled by base-pairing interactions formed in cis or trans , or by other binding proteins and the unwinding/remodeling activity of helicases. The actions of all these factors modulate substrate access.
In the degradation pathway of mRNA for E. coli , the initial cleavage of a transcript by RNase E is followed closely by exonucleolytic degradation of the products by PNPase (polynucleotide phosphorylase), RNase II, or RNase R ().
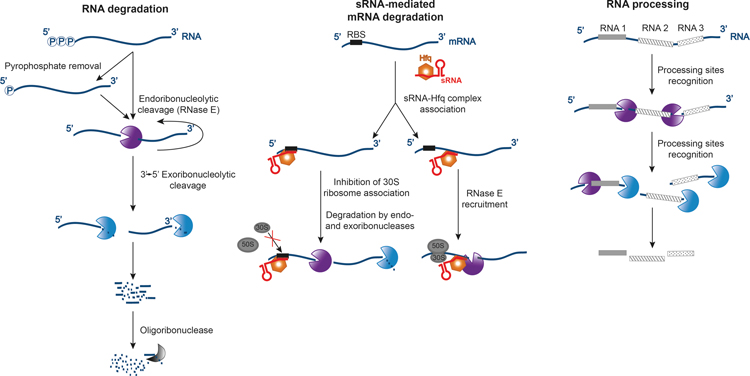
Figure 1: RNase-dependent processes in bacteria. RNases play crucial roles in efficient removal of defective or unnecessary RNAs, regulation of gene expression by sRNAs, and processing of various types of RNAs. (Left) RNA degradation is initiated by endoribonucleolytic cleavage, which can be preceded by pyrophosphate removal from the primary transcript. The majority of degradation initiation events are RNase E dependent. The initial cleavage generates monophosphorylated RNA fragments that can either boost subsequent RNase E cleavage or become substrates for cellular exoribonucleases. Fragments resulting from exoribonucleolytic degradation are further converted to nucleotides by oligoribonuclease. (Middle) When RNA degradation is mediated by sRNA, sRNA-chaperone complexes (such as sRNA-Hfq) can recognize a complementary sequence near the translation initiation region and prevent ribosome association on the transcript (left branch). Naked mRNA is rapidly scavenged by endo- and exoribonucleases. The sRNA-Hfq complex can also bind within the coding region of mRNA, recruiting RNase E and promoting transcript decay (right branch). (Right) In the case of substrates for processing, the order of RNA processing can be defined by the structure of precursors and the specificity of the RNases. The processing can form a cascade of interdependent events where some target sites are being revealed only upon specific initial cleavage. RNA, dark blue; endoribonucleases, purple; exoribonucleases, light blue; sRNA, red; ribosomes, gray ovals; Hfq, orange.
Font size:
Interval:
Bookmark:
Similar books «Regulating with RNA in Bacteria and Archaea»
Look at similar books to Regulating with RNA in Bacteria and Archaea. We have selected literature similar in name and meaning in the hope of providing readers with more options to find new, interesting, not yet read works.
Discussion, reviews of the book Regulating with RNA in Bacteria and Archaea and just readers' own opinions. Leave your comments, write what you think about the work, its meaning or the main characters. Specify what exactly you liked and what you didn't like, and why you think so.