Corien Bakermans - Microbial Evolution under Extreme Conditions
Here you can read online Corien Bakermans - Microbial Evolution under Extreme Conditions full text of the book (entire story) in english for free. Download pdf and epub, get meaning, cover and reviews about this ebook. year: 2015, publisher: De Gruyter, genre: Romance novel. Description of the work, (preface) as well as reviews are available. Best literature library LitArk.com created for fans of good reading and offers a wide selection of genres:
Romance novel
Science fiction
Adventure
Detective
Science
History
Home and family
Prose
Art
Politics
Computer
Non-fiction
Religion
Business
Children
Humor
Choose a favorite category and find really read worthwhile books. Enjoy immersion in the world of imagination, feel the emotions of the characters or learn something new for yourself, make an fascinating discovery.

- Book:Microbial Evolution under Extreme Conditions
- Author:
- Publisher:De Gruyter
- Genre:
- Year:2015
- Rating:3 / 5
- Favourites:Add to favourites
- Your mark:
Microbial Evolution under Extreme Conditions: summary, description and annotation
We offer to read an annotation, description, summary or preface (depends on what the author of the book "Microbial Evolution under Extreme Conditions" wrote himself). If you haven't found the necessary information about the book — write in the comments, we will try to find it.
Corien Bakermans: author's other books
Who wrote Microbial Evolution under Extreme Conditions? Find out the surname, the name of the author of the book and a list of all author's works by series.











Microbial Evolution under Extreme Conditions — read online for free the complete book (whole text) full work
Below is the text of the book, divided by pages. System saving the place of the last page read, allows you to conveniently read the book "Microbial Evolution under Extreme Conditions" online for free, without having to search again every time where you left off. Put a bookmark, and you can go to the page where you finished reading at any time.
Font size:
Interval:
Bookmark:

Corien Bakermans
Todays microorganisms represent the vast majority of biodiversity on Earth and have survived nearly 4 billion years of evolutionary change. Microbial evolution occurred and continues to take place in a vast variety of environmental conditions that range, for example, from anoxic to oxic, from hot to cold, and from free-living to symbiotic. Some of these physicochemical conditions are considered extreme, particularly when inhabitants are limited to microorganisms. It is easy to imagine that microbial life in extreme environments is somehow more constrained and perhaps subjected to different evolutionary pressures. But what do we actually know about microbial evolution under extreme conditions and how can the knowledge gained from extreme environments as model systems be applied to other conditions?
Extreme environments generally have physicochemical conditions that are challenging to cells and their macromolecules ( .) Environments with physicochemical extremes (i.e. pH or temperature) are considered in this volume, as well as environments with extremely low nutrient concentrations ( Chapter 7) and extraordinary cell-cell interactions (symbiosis, Chapter 12). Most of the extreme conditions examined occur in the natural (versus human created) world; however, anthropogenic environments such as acid mine drainage (AMD, Chapter 2) and extreme conditions that occur in our homes (such as extreme heat of dishwashers, extreme dryness, and variations therein) are included ( Chapter 10).
. Characteristics of extreme environments.
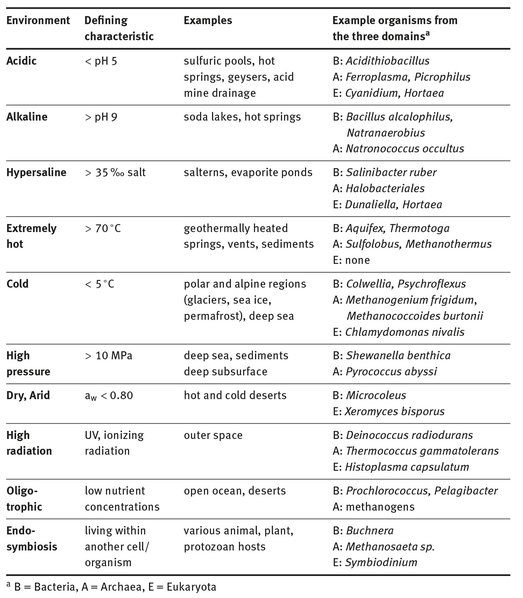
The organisms that inhabit extreme environments are called extremophiles because they thrive in the physicochemical conditions of extreme environments. These extremophiles can be found in all three domains (Archaea, Bacteria, and Eukaryota), although sometimes with a limited phylogenetic distribution. Some extremophiles are polyextremophiles ( Chapters 8 and 10) that thrive in multiple extreme conditions (e.g. high temperature, pH, and salt concentrations polyextremotolerant (i.e. tolerating, but not thriving in, the extreme conditions). While extremophiles succeed in extreme environments, the challenges presented by the physicochemical conditions of extreme environments commonly manifest as a lower abundance and diversity of inhabitants. For example, diversity decreases with increasing and Chapter 4).
With their relatively limited numbers of inhabitants, extreme environments can serve as good model systems for the study of evolutionary processes , helping to reduce the need for cultivation. Extreme environments often have fewer physicochemical variables in the sense that one condition (such as pH or temperature) may dominate, thereby reducing variability in the system.
Extreme environments are also good models seeing as their geographic boundaries, and often their geological history, can be mapped. The history and distribution of extreme environments will affect the evolutionary history of extremophiles (and their adaptive traits). For example, currently high temperature (hydrothermal) systems are relatively isolated from one another geographically, yet plate tectonics and volcanism have insured that high temperature environments, which likely dominated the early earth ( Chapter 9), have existed throughout earths history and alkaliphiles may be ancient and could have lots of interactions with other acidophiles and alkaliphiles .
All of these factors allow extreme environments to serve as model systems. Ecosystems with a dominant species or physicochemical condition can be very useful for disentangling the effects of different factors on the processes and mechanisms of microbial evolution. In particular, the variety of extreme environments, and especially comparisons between environments, will further facilitate the examination of microbial evolution and evolutionary processes across an entire spectrum of conditions from low to high diversity, low to high cell density, and low to high variability of physicochemical or biological conditions.
The study of evolution in microbes is complicated by their small size, rapid growth, asexual reproduction, horizontal gene transfer (HGT), living in close contact with diverse organisms, and the variability of their microhabitats. While an exhaustive review of microbial evolution is beyond the scope of this chapter, some key aspects of microbial evolution will be summarized here.
Our current understanding of microbial evolution suggests that it is complex and that there is no one evolutionary process that applies to and explains all microbes in all contexts (for current perspectives see . Regardless of whether trees or webs best represent the evolutionary history of prokaryotes, vertical descent sustained by binary fission is an important component of microbial evolution.
Evolution cannot ensue without genetic variation, which can be introduced into microbial cells by a variety of mechanisms that include replication errors, stress-induced mutations . HGT is a major contributor to genetic variation in prokaryotes . Genetic variation, however it is introduced, is fundamental to the ability of microorganisms to evolve.
Gene gain and loss are also important evolutionary processes in prokaryotes. Genome expansion can occur through the gain of genes via HGT (as discussed above), de novo gene creation, or gene duplication. Gene gain can be extensive as each of these processes may contribute up to 41% of the genes in prokaryotic genomes. De novo created genes (a.k.a. orphan genes or apparently contribute to the compact gene-rich genomes typical of prokaryotes.
Subsequently, natural selection can act on genetic variation to form clusters of genotypes that may represent species or ecotypes within communities. In positive selection traits that confer an advantage survive and increase within a population, while in negative (or purifying) selection traits that do not confer an advantage are removed from the population. Selective sweeps will promote retention of advantageous .
Measures of population and community structure are of course affected by the definition of microbial species (for current perspectives see . While microbiologists generally agree that there is a large amount of variability within microbial species, debate continues on how to delineate cutoffs in variability that correspond to species and how these cutoffs should relate to functional differences.
From the above summary, it is apparent that many variables, mechanisms, and processes are at work during microbial evolution contributing to its complexity. Reductionist studies in the laboratory contribute a great deal to our understanding of microbial evolution (e.g. along with a range of environments (extreme and otherwise) to sample, evolutionary microbiology is poised for significant advancement and synthesis in our understanding of how evolutionary and ecological processes (fine scale to large scale) determine the structure of microbial species and communities.
.
Many questions remain about how evolution versus ecology shapes microbial communities, particularly those in extreme environments. Ultimately, the relative rates of forces that increase variation (e.g. mutation, recombination ) and enough time, the diversity of extreme environments should approach that of non-extreme environments. Perhaps the low diversity of extreme environments is maintained by more frequent selective sweeps or bottlenecks possibly aided by relative temporal and spatial homogeneity of the environment and/or geographic isolation which limits immigration of extremophilic organisms and adaptive traits. Extreme environments, with their relatively low diversity, are good models for determining how ecology versus evolution shapes microbial communities.
Font size:
Interval:
Bookmark:
Similar books «Microbial Evolution under Extreme Conditions»
Look at similar books to Microbial Evolution under Extreme Conditions. We have selected literature similar in name and meaning in the hope of providing readers with more options to find new, interesting, not yet read works.
Discussion, reviews of the book Microbial Evolution under Extreme Conditions and just readers' own opinions. Leave your comments, write what you think about the work, its meaning or the main characters. Specify what exactly you liked and what you didn't like, and why you think so.