Hans-Georg Liebich - Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference)
Here you can read online Hans-Georg Liebich - Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference) full text of the book (entire story) in english for free. Download pdf and epub, get meaning, cover and reviews about this ebook. year: 2019, publisher: 5m Publishing, genre: Home and family. Description of the work, (preface) as well as reviews are available. Best literature library LitArk.com created for fans of good reading and offers a wide selection of genres:
Romance novel
Science fiction
Adventure
Detective
Science
History
Home and family
Prose
Art
Politics
Computer
Non-fiction
Religion
Business
Children
Humor
Choose a favorite category and find really read worthwhile books. Enjoy immersion in the world of imagination, feel the emotions of the characters or learn something new for yourself, make an fascinating discovery.

- Book:Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference)
- Author:
- Publisher:5m Publishing
- Genre:
- Year:2019
- Rating:4 / 5
- Favourites:Add to favourites
- Your mark:
Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference): summary, description and annotation
We offer to read an annotation, description, summary or preface (depends on what the author of the book "Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference)" wrote himself). If you haven't found the necessary information about the book — write in the comments, we will try to find it.
Hans-Georg Liebich: author's other books
Who wrote Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference)? Find out the surname, the name of the author of the book and a list of all author's works by series.






Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference) — read online for free the complete book (whole text) full work
Below is the text of the book, divided by pages. System saving the place of the last page read, allows you to conveniently read the book "Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference)" online for free, without having to search again every time where you left off. Put a bookmark, and you can go to the page where you finished reading at any time.
Font size:
Interval:
Bookmark:
The cell (cellula)
Eukaryotic cells contain a soluble matrix (cytosol) in which numerous organelles and inclusions are suspended. These cellular contents are referred to collectively as the cytoplasm. Cellular processes associated with metabolism, respiration, energy transformation, contractility and cell motility take place within the cytoplasm. These processes involve functional interactions between the cytosol and the organelles. The matrix is an aqueous gel incorporating variably sized molecules and structural elements in the form of a cytoskeleton.
Most organelles are bounded by a biological membrane that, by virtue of its specialised layered structure, separates the individual organelles from the surrounding matrix ( compartmentation ). This permits diverse metabolic processes to occur independently within the confines of the same cell. The cellular organelles can be subdivided as follows:
- membranous organelles:
- nucleus,
- endocytotic vesicles,
- exocytotic vesicles,
- endosomes,
- lysosomes,
- peroxisomes,
- rough endoplasmic reticulum (rER),
- smooth endoplasmic reticulum (sER),
- Golgi apparatus,
- mitochondria,
- non-membranous organelles:
- ribosomes,
- microtubules,
- filaments (actin filaments, intermediate filaments) and
- centrioles.
The nucleus directs all of the functions of the cell and houses its genetic material . Strands of deoxyribonucleic acid (DNA) within the nucleus store genetic information and serve as a regulatory and information-processing system for cellular metabolism. The intracellular organelles act synergistically, their combined actions giving rise to the specific functions of a particular cell. As these are clearly distinguishable under the light microscope, the cytoplasm and the nucleus are conventionally regarded as separate compartments. The nucleus is basophilic (= binds with basic stains, e.g. stains blue with haematoxylin), while the cytoplasm is predominantly acidophilic (binds with acidic stains, e.g. stains red with eosin). The cytoplasm is surrounded by a highly differentiated biological membrane, the plasma membrane ( plasmalemma ), that serves both to separate the cell from its surroundings and to establish a connection between the cell and its environment.
Cell membranes are biological membranes that constitute the structural foundation of most organelles and form the outer boundary of the cell. The outer (surface) membrane is highly specialised and is termed the plasmalemma.
All biological membranes have a similar basic structure ( unit membrane ) comprising layers of phospholipid and protein molecules held together by non-covalent bonds. Chains of lipid molecules form a lipid bilayer that encloses, or is traversed by, protein molecules and glycoproteins in a ratio of 2:1 (depending on cell type, this relationship varies from 4:1 to 1:4).
With a total thickness of 7.5 to 10 nm, the cell membrane can only be seen using electron microscopy. When prepared with osmic acid, the cell membrane appears as three layers: an electron-dense outer layer ( lamina externa ), an inner layer ( lamina interna ) and an electron-lucent intermediate layer ( lamina intermedia ).
On the outer surface of the plasmalemma, carbohydrates (oligosaccharide chains) attach to membrane proteins, forming glycoproteins, and to lipids of the bilayer, forming glycolipids. This gives rise to a layer known as the glycocalyx (cell coat).
The cell membrane is a biological membrane () comprising lipid-protein layers covered on their external surface by oligosaccharide chains. The individual components of the cell membrane comprise:
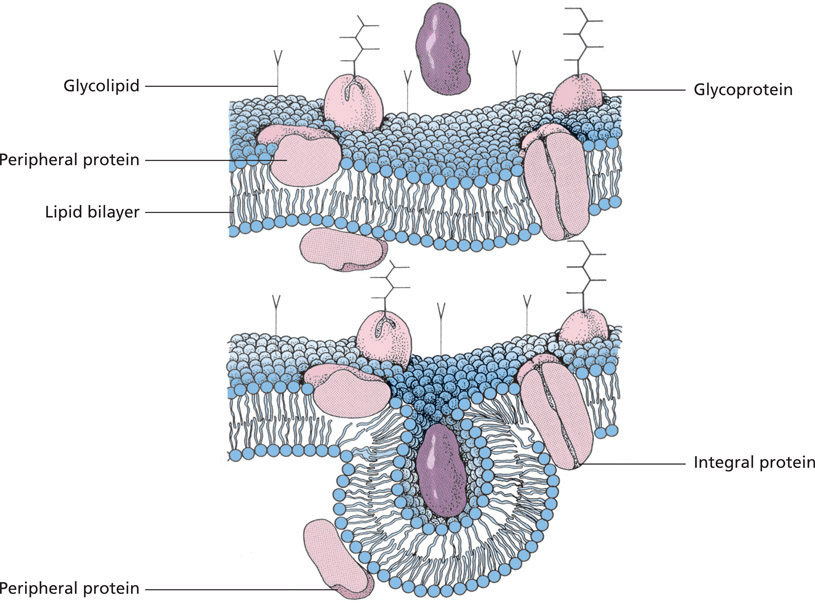
Structure of the cell membrane (schematic). The membrane consists of a lipid bilayer with associated membrane proteins. Carbohydrate side chains attach to the outer surface of the membrane, giving rise to the glycolipids and glycoproteins that make up the glycocalyx. During uptake of ligands (e.g. proteins), the membrane invaginates to form an endocytotic vesicle.
- a lipid bilayer,
- membrane proteins and
- surface polysaccharides.
The lipid molecules of the cell membrane are arranged in a double layer . Phospholipids are the main component of the bilayer, with cholesterol and glycolipids present in smaller quantities. The lipids are amphipathic, with one hydrophilic (affinity for water, polar), electrically charged head group and two hydrophobic (water-repelling, non-polar) hydrocarbon tails. The hydrophobic tails consist alternately of saturated and unsaturated fatty acids. The hydrophilic head groups are oriented towards the outer surfaces of the membrane while the hydrophobic tails face towards each other. Due to their specific lipid molecule content, and the concentration of electric charge in the outer membrane components, most membranes are asymmetrical ().
The structure of the cell membrane has important functional implications. The variable length of the saturated and unsaturated hydrocarbon chains, and their ability to move, influences the dynamics of the membrane, giving it a fluid quality . At a molecular level, the fluidity of the membrane is determined by the ability of the phospholipids to rotate and diffuse laterally within the membrane. Less frequently, they also flip-flop between individual layers of the bilayer.
The fluidity of the cell membrane gives rise to the plasticity of the cell as a whole. Components of the membrane may be pinched off (e.g. by exocytosis or endocytosis) or replaced. Following interruption by external influences or cellular secretory activity, the lipid bilayer is re-established spontaneously within the aqueous microenvironment, thus restoring the continuity of the membrane.
Cholesterol is an important functional component of the membrane, serving to regulate the intramembranous movement of lipid chains and to increase the mechanical strength of the bilayer .
Glycolipids are present in the outer layer of the plasmalemma of all animal cells and contribute significantly to the asymmetry of the cell membrane. Their head groups, consisting of one or more polar sugar residues (oligosaccharides), project beyond the surface of the membrane. Glycolipids consist predominantly of neutral glycolipids (e.g. galactocerebroside within myelin sheaths) or gangliosides (e.g. sialic acids in cell receptors) (for further information refer to Membrane polysaccharides, below, and biochemistry texts).
The proteins of the cell membrane are incorporated into the lipid bilayer, where they perform membrane-specific functions (). Membrane proteins include transport proteins, enzymes and specific receptor proteins. These are globular and, like the membrane lipids, amphipathic. Membrane proteins similarly exhibit hydro-phobic inwardly oriented regions and hydrophilic moieties that project from one side or, more commonly, both sides of the membrane.
Several types of membrane proteins are recognised. These are named according to their functions:
- pumps: e.g. for amino acids, sugars, Na+, Ca2+,
- channel proteins: passive diffusion of ions or molecules through the membrane,
- receptor proteins: cell recognition and binding of ligands (e.g. hormones, antibody reactions),
- linking proteins: binding with the intracellular cyto-skeleton (e.g. actin filaments) or the extracellular matrix (e.g. fibronectins),
- enzymes: e.g. ATPase associated with the inner membrane of mitochondria, disaccharidases and dipeptidases involved in digestion and
Font size:
Interval:
Bookmark:
Similar books «Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference)»
Look at similar books to Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference). We have selected literature similar in name and meaning in the hope of providing readers with more options to find new, interesting, not yet read works.
Discussion, reviews of the book Veterinary Histology of Domestic Mammals and Birds 5th Edition: Textbook and Colour Atlas (Veterinary Clinical Reference) and just readers' own opinions. Leave your comments, write what you think about the work, its meaning or the main characters. Specify what exactly you liked and what you didn't like, and why you think so.